Click here if you are looking for a certified online tutor or home tutor

Reproductive System
| I | INTRODUCTION |
Reproductive
System, term applied to the group of plant or animal organs that are
necessary for or that are accessory to the reproductive processes (see
Reproduction). The basic units of sexual reproduction are the male and
female germ cells; this article deals with the organs within which the germ
cells of animals mature and are stored, the organs through which they are
transported in the process of producing a new individual, and accessory
glandular organs. For the reproductive organs of plants, see Plant
Propagation.
| II | ORIGIN OF THE REPRODUCTIVE CELLS |
When the embryo of any sexually reproducing
animal is undergoing cell division, certain cells, known as primordial germ
cells, which are produced by such division, remain in an undifferentiated state.
Cells other than primordial germ cells are known as vegetative cells or somatic
cells; these cells become differentiated into tissues and organs. In
invertebrates, the primordial germ cells congregate in the body cavity or in a
section of the circulatory system; in vertebrates, these cells are located in
organs that adjoin the excretory system. The tissues in which the germ cells
lodge become reproductive organs known as gonads. These organs are
embryologically derived from primitive kidneys located in the anterior, lateral
part of the body; in most mammals, they shift before birth to the posterior,
ventral region of the body. The primordial germ cells remain inactive in the
gonads until the animal reaches sexual maturity, when the undifferentiated cells
undergo a great number of normal cell divisions or mitoses. In the process of
developing into mature reproductive cells or gametes, the germ cells undergo a
special type of cell division, known as meiosis, which halves the number of
chromosomes they carry. At the time of sexual maturity, the somatic cells
composing the gonads of higher animals begin to secrete hormones that control
the appearance of the various secondary sexual characteristics (see
Sex).
| III | GONADS |
Organs that contain germ cells which later
develop into male gametes or spermatozoa are known as testes or male gonads.
Organs that contain germ cells which later develop into female gametes, eggs, or
ova are known as ovaries. In many invertebrate species individual animals bear
both testes and ovaries (see Hermaphroditism). In some invertebrates, and
in most vertebrates, individuals bear either testes or ovaries, but not both
sets of organs. In invertebrates, a single animal may have as many as 26 pairs
of gonads; in vertebrates, the usual number is 2. Cyclostomes and most birds are
unusual among vertebrates in possessing only a single gonad; owls, pigeons,
hawks, and parrots are unusual among birds in having two gonads. The size of
gonads increases at sexual maturity because of the great number of germ cells
produced at that time; many germ cells are also produced during breeding seasons
so that many animals have a seasonal increase in size of the gonads. During the
breeding season of fish, the ovaries increase in size until they constitute
about one-quarter to one-third of the total body weight.
The testes and ovaries of mature animals
differ greatly in structure. The testes are composed of delicate convoluted
tubules, known as seminiferous tubules, in which the primitive germ cells mature
into spermatozoa. The testes of mammals are generally oval bodies, enclosed by a
capsule of tough connective tissue. Projections from this tough capsule into the
testis divide the testis into several compartments, each of which is filled with
hundreds of seminiferous tubules. The mature spermatozoa are discharged through
a number of ducts, called the efferent ducts, which communicate with the
epididymis, a thick-walled, coiled duct in which the sperm are stored.
In all vertebrates below marsupials on the
zoological scale, and in elephants, sea cows, and whales, the testis remains
within the body cavity during the lifetime of the animals. In many mammals, such
as rodents, bats, and members of the camel family, the testis remains within the
body cavity during periods of quiescence, but moves into an external pocket of
skin and muscle, known as the scrotum, during the breeding season. In
marsupials, and in most higher mammals, including the human male, the testes are
always enclosed in an external scrotum. During fetal life, the testes move
through the muscles composing the posterior, ventral portion of the trunk and
carry with them the portion of the peritoneum and skin surrounding these
muscles. The channel in the muscles through which the testis moves is known as
the inguinal canal; it usually closes after birth, but sometimes remains open
and is then often the site of herniation (see Hernia). The portion of the
peritoneum that the testis carries with it forms a double wall of membrane
between the scrotum and testis and is known as the tunica vaginalis.
Occasionally, the testes in the human male do not descend into the scrotal sac;
this condition of nondescent, which is known as cryptorchidism, may result in
sterility if not corrected by surgery or the administration of hormones.
Retention of the testes within the body cavity subjects the germ cells to
temperatures that are too high for their normal development; the descent of the
testes into the scrotum in higher animals keeps the testes at optimum
temperatures.
Unlike germ cells in the testis, female germ
cells originate as single cells in the embryonic tissue that later develops into
an ovary. At maturity, after the production of ova from the female germ cells,
groups of ovary cells surrounding each ovum develop into “follicle cells” that
secrete nutriment for the contained egg. As the ovum is prepared for release
during the breeding season, the tissue surrounding the ovum hollows out and
becomes filled with fluid and at the same time moves to the surface of the
ovary; this mass of tissue, fluid, and ovum is known as a Graafian follicle. The
ovary of the adult is merely a mass of glandular and connective tissue
containing numerous Graafian follicles at various stages of maturity. When the
Graafian follicle is completely mature, it bursts through the surface of the
ovary, releasing the ovum, which is then ready for fertilization; the release of
the ovum from the ovary is known as ovulation. The space formerly occupied by
the Graafian follicle is filled by a blood clot known as the corpus
hemorrhagicum; in four or five days this clot is replaced by a mass of yellow
cells known as the corpus luteum, which secretes hormones playing an important
part in preparation of the uterus for the reception of a fertilized ovum. If the
ovum goes unfertilized, the corpus luteum is eventually replaced by scar tissue
known as the corpus albicans. The ovary is located in the body cavity, attached
to the peritoneum that lines this cavity.
The functioning of both male and female
gonads is under the hormonal influence of the pituitary gland.
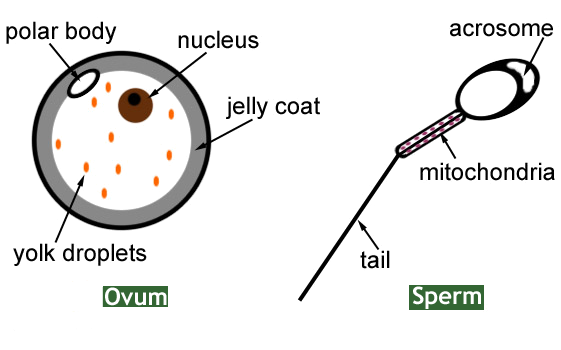 |
| Ovum and Sperm |
| IV | TRANSPORTATION OF THE REPRODUCTIVE CELLS |
Before being discharged from the body, the
reproductive cells travel from the gonads to an external body opening. In many
invertebrates, and in a few aquatic vertebrates, the reproductive cells are
discharged into water directly from the gonads through pores in the body wall.
In higher animals ducts carry the reproductive cells into the urinary or cloacal
excretory systems, or into independent reproductive passages.
In male vertebrates, the ducts are directly
connected to the testes. The male ducts include the epididymis, which lies
attached to the testis, and which transports the male gametes to the vas
deferens. The vas deferens carries the spermatozoa to the ejaculatory duct, the
contractions of which cause the discharge of sperm into the posterior
urethra.
In most fishes, the ovary has a hollow
expansion through which the ova pass into the cloaca. In most other vertebrates,
however, no direct connection exists between the ovary and the oviducts that
carry the ova into the cloaca or into the independent external opening. In
mammals, when the Graafian follicle bursts, the egg falls toward the interior of
the abdominal cavity. The oviduct (known in higher mammals as the fallopian
tube) has an open, funnel-shaped end located near the ovary, and the mature egg
is drawn into the funnel by ciliary action. Occasionally, the egg misses the
open end of the oviduct and falls into the abdominal cavity; such eggs are
capable of being fertilized, resulting in ectopic pregnancies (see
Pregnancy and Childbirth). In animals lower than marsupials the oviducts
open directly into the cloaca; in marsupials and placental mammals, the
oviducts, two of which are normally present, fuse at their cloacal ends to form
a thick, muscular organ, the uterus or womb, in which the young develop, and a
thinner channel, the vagina, which opens exteriorly.
 |
| sperm, ova |
| V | GENITALS |
In animals that lay their eggs and discharge
their sperm into water, the spermatozoa reach the eggs by chemical attraction;
the eggs of individuals of a species attract only the sperm of members of the
same species. When eggs and sperm are deposited at great distances from each
other, the number of eggs fertilized is small. Many amphibians and aquatic
vertebrates solve this problem by attaching themselves to their mates by means
of holdfast mechanisms, such as claspers; when the female of these animals
deposits the eggs, the male can immediately deposit sperm in the same
location.
In terrestrial animals, various adaptations
have been developed whereby internal fertilization of the eggs may be produced.
The male snake, which discharges the spermatozoa through a cloaca, has, for
example, anal hooks that are inserted into the cloaca of the female during the
breeding season. These hooks bind the male and the female together while the
spermatozoa are being discharged. External reproductive organs used by animals
to facilitate internal fertilization are known as genitals or genitalia.
The male genital of all mammals above the
monotremes is the penis, a protrusible, erectile organ that directs the
spermatozoa into the female cloaca or vagina. In turtles and crocodiles, which
are the most primitive animals possessing the organ, the penis is located on the
ventral wall of the cloaca and is grooved along its upper side. The spermatozoa
travel along the groove into the female cloaca. In marsupials and placental
mammals, the penis is a closed tube, composed of three bundles of tissue, bound
together by connective tissue and covered with loose skin. Two large bundles of
tissue, known as the corpora cavernosa, constitute the upper portion of the
penis; these bundles contain numerous compartments that are filled with blood
during sexual excitement, causing the penis to become stiff and erect. The flow
of blood into the corpora cavernosa is controlled by sacral spinal nerves. Below
and between the corpora cavernosa is the third bundle of tissue, which is known
as the corpus spongiosum; this bundle is perforated by the urethra and, in
several lower mammals, also contains a bone that serves to further stiffen the
male genital. At the terminal end of the penis is a sensitive cap known as the
glans; in marsupials the glans is forked. In many mammals, the male genital,
when not erect, is withdrawn into a sheath in the body. In primates, including
the human male, the penis is pendant, and the glans is covered with a layer of
retractile skin, known as the prepuce or foreskin, which corresponds to the
sheath of lower animals.
The chief female genital is known as the
vagina. This organ is present in all marsupials and placental mammals. In
marsupials, two vaginas and two uteri are present; in primates, including the
human female, only one vagina is present; and in mammals intermediate in
development between the marsupials and primates, various degrees of partially
united, double vaginas are present. The external end of the vagina is covered in
virgin primates by a membrane, known as the hymen. Anterior to the hymen is the
external opening of the urethra. The urethra and the opening of the vagina are
contained in an indented space which is known as the vestibule. In primates, two
membranous folds, known as the labia minora, are found on either side of the
vestibule. In primates, including the human female, two additional folds, the
labia majora, enclose the labia minora. The clitoris, at the front of the labia,
is homologous with the penis although greatly reduced in size (see Homology
below).
| VI | ACCESSORY GLANDS |
Among the glands accessory to the
reproductive process are those that provide a fluid medium in which the
spermatozoa may live, those that produce mucus which reduces friction during
copulation, those that emit alluring odors to members of the opposite sex, and
those that secrete nourishment for the ova, the embryos, and the newborn
young.
The seminal vesicles of the male have already
been mentioned as organs that secrete mucus. The most important male accessory
gland is the prostate gland, a compound gland about the size of a chestnut
located at the base of the urethra where the urethra leaves the bladder and
enters the penis. The prostate secretes a thin milky fluid with a characteristic
odor; this fluid constitutes the greater part of the semen that is deposited in
the female vagina and that contains the spermatozoa. The prostate gland is
present only in placental mammals, and among these mammals is absent in
edentates, martens, otters, and badgers. Cowper's glands are two small glands,
about the size of a pea, located on each side of the base of the penis. Their
secretion is thick and clear and is believed to protect the spermatozoa against
excess vaginal acidity. Cowper's glands are absent in bears, dogs, and aquatic
mammals.
The primary lubricating glands of the female
are the cervical glands, located in the uterus where the uterus communicates
with the vagina, and Bartholin's glands, located in the vestibule between the
hymen and the labia minora. Both sets of glands secrete mucus. Female placental
mammals also have uterine glands that prepare the uterus for the reception of
the fertilized egg.
The anal glands of many mammals secrete
special substances called pheromones, which signal reproductive readiness by
scent to members of the opposite sex. Pheromones may occur in other glandular
secretions as well.
Among the different structures serving to
nourish the young, the placenta of placental mammals is unique (see
Fetus). The mammary glands of mammals are also included among accessory
reproductive glands (see Breast). Female egg-laying animals have albumen
glands, which coat the zygote with nutrient albumen before the egg is laid, and
nidamental glands, which surround the zygote and albumen with a leathery or
calcareous shell.
| VII | HOMOLOGY |
The sex of a young embryo is
indistinguishable, males and females passing through similar embryonic stages.
Embryo males and females develop almost a duplicate set of reproductive organs,
one set becoming vestigal shortly before birth, and the other set becoming
prominent. Most cases of so-called mammalian hermaphroditism are actually cases
of abnormal development in which external genitalia similar to those of both
sexes have been developed. For example, mammalian females have a small, erectile
organ, consisting of two corpora cavernosa, located in the upper portion of the
vestibule. This organ, called the clitoris, is homologous with the male penis;
except in lemurs and a few rodents it does not contain the urethra, which is
usually located beneath the clitoris. In species in which the male possesses a
penis bone, the clitoris of the female contains a small bone.
Menstrual Cycle
 |
| Menstruation An average menstrual cycle begins with three to five days of menstruation, the shedding of the uterine lining, during which hormone levels are low. At the end of menstruation, a pituitary hormone stimulates new follicles to develop in the ovary. These secrete estrogen as they mature, causing cells in the lining of the uterus to proliferate. Mid-cycle, one mature follicle releases an egg. The empty follicle forms the corpus luteum, an endocrine body that secretes progesterone. Under the added influence of progesterone, the uterine lining thickens further and swells in preparation for the implantation of a fertilized egg. If fertilization does not take place, the corpus luteum dies and hormone levels fall. Without hormonal support, the uterine lining disintegrates and discharges, beginning a new menstrual period and cycle. |
Menstruation, periodic
vaginal discharge in humans and other mammals, consisting of blood and cells
shed from the endometrium, or lining of the uterus (see Reproductive
System). Menstruation accompanies a woman's childbearing years, usually
beginning between the ages of 10 and 16, at puberty, and most often ceasing
between the ages of 45 and 50, at menopause. Menstruation is part of the process
that prepares a woman for pregnancy. Each month the lining of the uterus
thickens; if pregnancy does not occur, this lining breaks down and is discharged
through the vagina. The three to seven days that menstruation lasts is called
the menstrual period.
In most women the menstrual cycle is about 28 days, but it
can vary considerably even from one month to another. The cycle is initiated by
hormones in the blood that stimulate the ovaries (the two female organs that
produce ova, or eggs). Each month, hormones cause an egg in one of the two
ovaries to mature (to become capable of being fertilized and develop into a
fetus). The ovaries also produce hormones of their own, primarily estrogen,
which cause the endometrium to thicken. About midway through the menstrual
cycle, 14 to 15 days before the next period, the ovary releases the mature egg
in a process called ovulation. The egg passes through the fallopian tube to the
uterus. If the egg unites with a sperm on its way to the uterus, fertilization
occurs and pregnancy ensues.
The three to five days the egg takes to reach the uterus
after being released by the ovary is known as the woman's fertile period. If
fertilization does occur, the fertilized egg attaches itself to the enriched
uterine lining and pregnancy continues. Menstruation does not occur during
pregnancy, and a missed period is often the first indication of pregnancy a
woman notices (see Pregnancy and Childbirth). If fertilization does not
occur, the lining of the uterus does not receive the hormones it needs to
continue the thickening process. Thus, the uterine lining breaks down and is
discharged from the body during menstruation.
Many women experience premenstrual discomfort. Tenderness
of the breasts and a tendency to retain fluid (bloat) are common one to seven
days before each period. Some women also experience a condition called
premenstrual syndrome (PMS), which is characterized by headaches, irritability,
nervousness, fatigue, crying spells, and depression with no apparent cause. A
few women also experience menstrual cramps (dysmenorrhea) during the first day
or two of the period. Although premenstrual symptoms and discomfort during
menstruation were once thought to be of psychological origin, research now
indicates that hormonal and chemical changes are responsible. New medications
are effective in treating these problems.

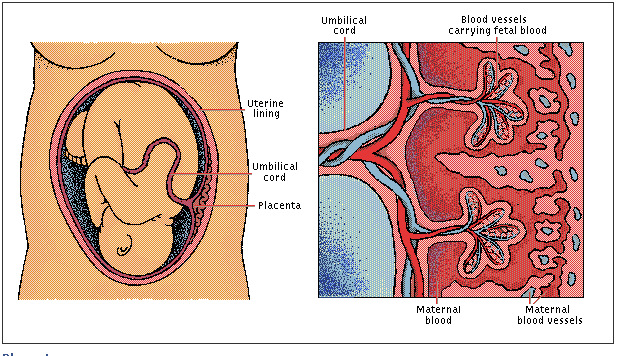 |
| Placenta and Umbilical Cord
Placenta
The placenta is responsible for
respiration and excretion in the growing fetus. Fetal blood flows through the
blood vessels of the umbilical cord to the placenta, where fingerlike
capillary nets are surrounded by pools of the mother’s blood. Here carbon
dioxide and other metabolic wastes diffuse from fetus to mother, and oxygen and
nutrients pass from mother to fetus. Some STDs can be passed from an infected
mother to her unborn child when disease-causing organisms cross the
placenta.
|